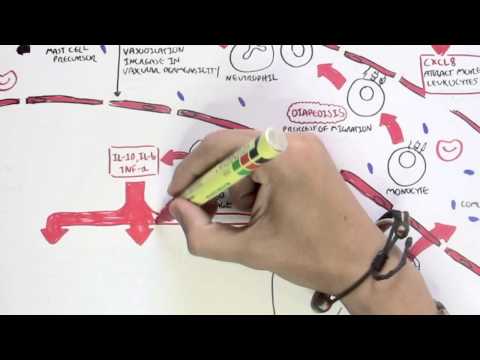
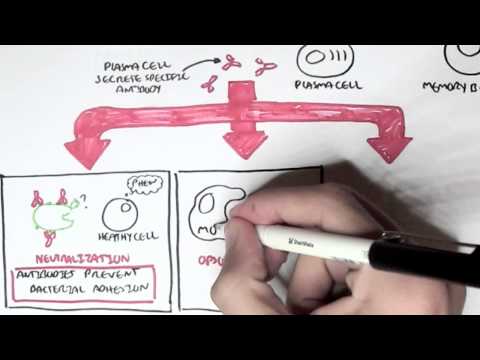
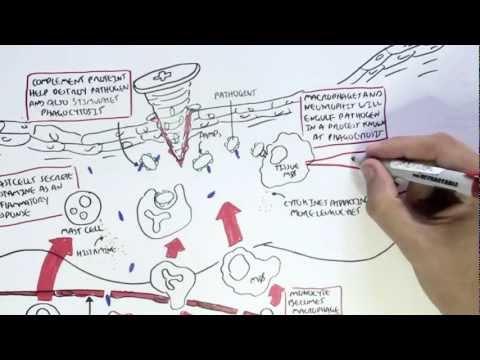
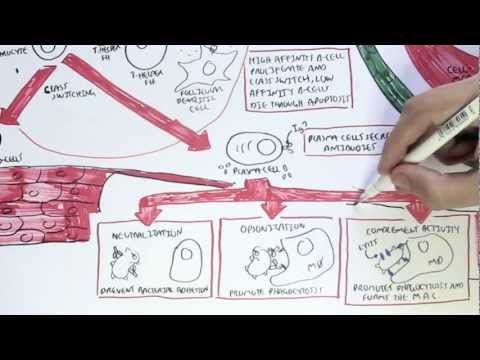
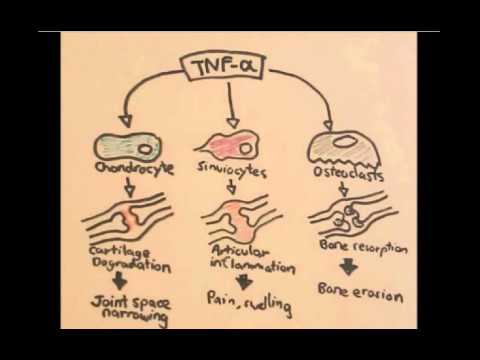
0:00 Arman Dohasudugan, Biology and Medicine videos, please make sure to subscribe, 0:05 join the form 0:06 and group for the latest videos, please visit Facebook, Arman Dohasudugan, like 0:10 , please. 0:11 And here you can also ask questions, answer some questions, and please post 0:14 some of your 0:15 artworks or any artworks, which you think are good. 0:19 And please change the quality settings to the highest one for better graphics. 0:23 In this video, we're going to talk about the complement system, and this is 0:25 just going 0:26 to be an overview to see how the complement proteins and the pathways are all 0:31 related 0:32 in some way. 0:34 Now what happens when a pathogen invades the body? 0:36 When the pathogen invades the body, the first line of defense is the innate 0:39 immune system, 0:40 which comprises the physical barriers and the chemical and biological barriers 0:44 afterwards. 0:45 Now when the pathogen successfully infiltrates our body, we have complement 0:50 proteins in the 0:51 bloodstream, the blood vessels, and these complement proteins are part of the 0:54 innate immune system. 0:56 And they are in the bloodstream because they're soluble proteins. 1:00 And what these complement proteins do, they do few things, including performing 1:05 obstinization, 1:06 making a membrane attack complex, and they also enhance inflammation. 1:12 Overall, the complement proteins aim in destroying the invading pathogen. 1:16 So what does obstinization mean? 1:18 Well, obstinization is a process where the complement proteins essentially 1:22 coats the outer 1:23 surface of the pathogen, which allows phagocytes, in this case, a macrophage to 1:28 engulf the pathogen 1:29 much more easier because the macrophage contains special receptors for specific 1:34 complement proteins. 1:36 Complement proteins can also make a membrane attack complex. 1:41 A membrane attack complex is essentially when a group of complement proteins 1:45 makes a hole 1:46 in a pathogen, which causes in-rushing fluids, creating a disbalance in osmolar 1:51 ity, which 1:52 causes the pathogen to lice, to be destroyed. 1:55 And we will see later on in this video how the complement proteins initiate 2:00 obstinization, 2:00 the membrane attack complex, and enhance inflammation. 2:03 But for now, let's see where complement proteins come from. 2:07 The complement system is composed of more than 30 types of proteins. 2:12 Now, the complement proteins is produced by the liver, and the complement 2:16 proteins will 2:16 then travel through the bloodstream. 2:18 Now, these complement proteins circulate in an inactive form, and so they don't 2:23 attack 2:23 the body, but when they come across a pathogen, they will become activated. 2:30 And when the complement proteins are activated, they will then obstinize, 2:34 create a membrane 2:34 attack complex on the pathogen, and also enhance inflammation. 2:39 The complement proteins don't actually get activated that easily, because 2:43 actually the 2:43 complement proteins interact with each other and induce a cascade of activation 2:48 , a complement 2:49 protein activating another complement protein. 2:53 And so because of this, there are many complement pathways, because there's 2:57 many types of ways 2:58 that complement proteins can activate each other. 3:02 And these complement pathways are the classical pathway, the alternative 3:05 pathway, and the 3:06 lectin pathway, the classical pathway being the first to be discovered. 3:11 The couple of proteins involved in the classical pathway is a C1Q, C1R, C1S, C4 3:17 , and C2. 3:18 In the alternative pathway, we have factor D, factor B, prepared in, and C3. 3:24 And for the lectin pathway, we have MBL, manos, bindin, lectin, fic colon, MASP 3:37 2, C4, and C2. 3:38 Now honestly, don't be intimidated by all these types of proteins, we will 3:41 learn about 3:42 them slowly. 3:43 But what is to know about this small diagram is that the classical pathway, the 3:47 alternative 3:48 pathway and the lectin pathway, all these proteins will essentially help in 3:53 splitting or activating 3:55 complement protein 3 to make a C3A and C3B. 4:01 And it is C3A, which enhances inflammation together with other common proteins, 4:06 and C3B, 4:07 which initiates obstinization, and as well as lysis of the cell to create a 4:12 membrane 4:12 attack complex. 4:15 So C3 is an important protein to remember, and C3 gets activated into 4:21 complement 3A and 4:22 complement 3B. 4:24 Now so complement proteins typically have a C in front of them. 4:29 So for example, we have C1, C2, and C3 here. 4:34 C2A proteins also tend to be activated when they are cleaved by something. 4:38 So for example, when C3 is cleaved to C3A and C3B, these are the activated 4:42 complement 4:42 proteins. 4:43 C2A can become activated to C2A and C2B. 4:47 C5 is cleaved to the active form of C5A and C5B. 4:53 I hope you understood that, but for now let's look at the complement pathways 4:57 and see how 4:58 they, all these pathways, will essentially activate C3 to cleave it into C3A 5:05 and C3B. 5:06 So remember, we have three complement pathways. 5:09 The classical pathway, the autone, or alternative pathway, and the lectin 5:13 pathway. 5:13 Here we have a pathogen, the screen. 5:16 And on the pathogen surface, we can either have antigens, we can have manos, 5:21 carbohydrates, 5:23 and also short own doseccharides, depending if it's a gram-negative or gram- 5:26 positive bacteria, 5:27 or whatever type of pathogen it is. 5:31 Let's look at the classical pathway first. 5:33 The classical pathway gets initiated or activated when antibodies bind to the 5:37 antigen of a pathogen. 5:39 And then when this happens, some complement proteins, composed of C1Q, C1S, and 5:44 C1R, bind 5:45 to these antibodies, will actually C1Q binds to the FC portion of the antibody. 5:52 And essentially what the C1S, C1Q, C1R complex does is that it forms another 5:58 complement protein 6:00 complex, known as the C4B2A complex. 6:05 Another name for this is the C3 convertase. 6:09 Because it is a C3 convertase, it is also an enzyme. 6:14 And we'll look at the function of C3 convertase later on. 6:17 But for now, let's stop there and look at the lectin pathway, which is very 6:20 much similar 6:21 to the classical pathway, but involves different types of proteins in the 6:25 activation of the 6:26 C4B2A complex, the C3 convertase. 6:29 Now the lectin pathway differs to the other pathways because what initiates it 6:34 are proteins 6:35 binding to carbohydrates on the pathogen. 6:38 So for example, we have a protein called Philcolin, which binds to the oligosac 6:42 charides of a pathogen, 6:44 a bacteria. 6:45 Philcolin also has other proteins bound to it, the MASP1 and MASP2. 6:51 Another type of protein is the manose binding lectin, or abbreviated MBL, which 6:56 binds to 6:57 manose parts of the pathogen. 7:04 Manose binding lectin also comprises of other proteins, the MASP1 and MASP2, 7:09 similar to 7:10 the Philcolin. 7:11 And MASP stands for essentially manose-associated serine protease. 7:15 So there's two types, the manose-associated serine protease 1 and manose- 7:18 associated serine 7:19 protease 2. 7:21 Now essentially what Philcolin and manose binding lectin do with the other man 7:25 ose-associated 7:27 serine protease is that they also form a complement protein complex, exactly 7:34 the same 7:35 as the classical one, the C4B2A complex, also known as C3 convertase. 7:42 So as you can see the classical pathway and lectin pathway involves different 7:47 proteins 7:47 in the initiation or the formation of C3 convertase known as the C4B2A complex. 7:56 And we skip the alternative pathway for now because the alternative pathway 7:59 actually becomes 7:59 activated when the classical pathway and lectin pathway forms this C4B2A 8:06 complex, the 8:07 C3 convertase, and we'll soon see why. 8:16 So now the question is what does the C4B2A complex do, this C3 convertase? 8:22 Well the C3 convertase is on the surface of the pathogen and what it 8:26 essentially does 8:26 is that it activates the C3 protein. 8:30 So the C4B2A complex splits the C3 to form C3B and C3A. 8:39 Now that we've got this formation of C3B and C3A, we go back to the alternative 8:44 pathway 8:45 and see how the alternative pathway is initiated. 8:49 The alternative pathway is initiated when the C4B2A convertase activates C3B. 8:56 The C3B then binds on the surface of the pathogen. 9:00 The alternative pathway will then form another type of C3 convertase different 9:06 to the ones 9:07 formed by the lectin and classical pathway. 9:10 This C3 convertase is called C3B-B-B complex. 9:18 The alternative pathway can also form another type of C3 convertase, this time 9:24 with a protein 9:25 called Propareden. 9:28 And this complex is known by the same name C3B-B-B complex, which is as I said 9:35 a C3 convertase. 9:38 So as you can see, the alternative pathway essentially enhances the classical 9:42 and lectin 9:43 pathway, or the classical and lectin pathway enhances the alternative pathway. 9:49 But all these pathways, the classical pathway, the alternative pathway and the 9:52 lectin pathway 9:53 all form a C3 convertase. 9:56 And what a C3 convertase does is that it activates the C3, which basically 10:01 splits a C3 10:03 into a C3A and a C3B. 10:07 So this is the main thing to take in. 10:13 Okay so now we have the activated form of C3, C3A and C3B. 10:16 Now what do they do? 10:18 C3A with other complement proteins such as C5A, which is actually one of the 10:21 most important 10:22 complement proteins, will enhance inflammation. 10:26 Actually all the complement proteins with A will do some form of inflammatory 10:31 process. 10:32 So in this case, C3A and C5A will stimulate mast cells to secrete histamine. 10:37 What does histamine do? 10:38 Histamine enhances inflammation. 10:40 Histamine increases vascular permeability, which allows leukocytes to pass 10:43 through more 10:44 easily. 10:45 And histamine also attracts leukocytes. 10:48 And in this case we can see a macrophage and neutrophil passing through. 10:53 Now what does C3B do? 10:55 And the important thing to know is that C3B can form Theo S. Gebont. 11:00 And C3B, if you remember at the beginning of the video, actually initiates obst 11:04 inization 11:05 and forms the initiates of the formation of the membrane attack complex. 11:10 Actually important thing to know is that there are actually many C3Bs and many 11:14 C3As 11:15 because the classical pathway, the alternative pathway and the electron pathway 11:18 , forms many 11:19 C3 convertase, which means that they split many C3s to form many C3Bs and many 11:27 C3As. 11:28 So it's important to know. 11:30 Now that we have many C3Bs, these C3Bs can actually bind to the pathogen 11:36 surface through 11:38 this Theo S. Gebont. 11:43 Now as you can see, we can see many C3Bs forming Theo S. Gebonts with pathogen 11:47 surface. 11:47 So it essentially coats the pathogen, a process called obstinization. 11:51 And this allows macrophage to easily engulf this pathogen once it's been coated 11:56 . 11:57 Now a macrophage actually has two special receptors for complement proteins, a 12:01 CR1 and 12:02 a C5A receptor. 12:04 A macrophage cannot engulf a pathogen coated with C3B just like that. 12:10 It actually needs the help of another protein, complement protein, called C5A. 12:16 So when C5A, complement protein, binds to the C5A receptor of the macrophage, 12:21 this then 12:22 allows the macrophage, or helps the macrophage, bind to the C3B protein through 12:29 the CR1 receptors. 12:31 And then through this, the macrophage can perform phagocytosis, eating up the 12:39 pathogen. 12:41 C3B can alternatively bind to the C3 convertase, specifically the C4B2A complex 12:48 from the classical 12:50 pathway or the lectin pathway, if you remember. 12:54 Now when it binds, the C3B will bind to this C4B2A complex, which will then 13:02 form the C4B2A3B 13:04 complex, also known as the C3C5 convertase. 13:10 And so this C3C5 convertase can do two things. 13:14 It can actually still split or activate C3 to form C3A and C3B. 13:22 And now it can actually cleave and activate C5 to C5A and C5B. 13:29 And remember, C5A can enhance inflammation, or it can bind to the macrophage's 13:35 receptor. 13:36 C5B is an important protein because it initiates the terminal stage, the 13:41 formation of the membrane 13:42 attack complex, abbreviated MAC, and so C5B together with other complement 13:51 proteins of 13:53 higher number, such as C9, C8, and C7, will form this mac, the membrane attack 14:00 complex. 14:01 And the formation of the membrane attack complex on a pathogen's surface 14:06 membrane will cause 14:08 the pathogen to lice to self-destruct, so cell bursting, destroying the path 14:14 ogen. 14:15 So as you can see from this overview of the complement system, the complement 14:20 proteins 14:20 and the system itself is very important for the body, especially in the first 14:26 line of 14:26 defense, in the innate immune system. 14:29 Because there was simply an overview of the complement system, I'll provide 14:32 links on the 14:32 screen which you can click to for the classical pathway, the alternative 14:35 pathway, and the 14:35 selecting pathway to see in more detail, as well as a link to the formation of 14:40 the membrane 14:40 attack complex, how it is formed. 14:42 So I hope you actually enjoyed this video. 14:44 Please like, comment, share, and please subscribe. 14:46 Thank you very much.